大脑这个结构精妙又交错纵横的器官一直以来是研究者们关心的研究对象,通过研究让我们的大脑来理解大脑,这一旅程想必十分有趣。而随着神经科学研究的不断深入,与之并行的是类脑科学的日新月异,除了想办法理解脑,研究者们还致力于如何模拟脑。为了向读者介绍如何从类脑科学的角度出发来理解大脑的工作机制,笔者计划通过“类脑”专栏和读者一起感受大脑的“机械”之美。
封面:19世纪机械娃娃工艺品草图
得益于当时工匠的奇技淫巧,勒内·笛卡尔(René Descartes)在1662年出版的L’homme《论人》中描写了这样一段经历:他在青年时期曾在以机械雕像闻名的圣杰曼·昂·雷(Saint-Germain-en-Laye)皇家花园散步,在园中看到一个栩栩如生的机器人跟他打招呼。仔细追问之下,工匠给他展示了机器人的内部结构。这个机器人由液压控制,打开对应的阀门,就会有水注入,让机器人做出相应的动作。
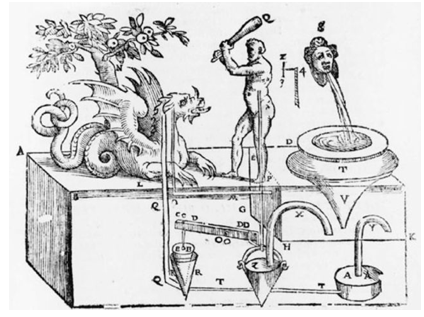
机械剧场
由亚历山大的Hero发明,该图片收录在其编写的Hero's Pneumatics的意大利译本中。
这个事件启发了笛卡尔,他认为“通过物质过程来解释生命是有可能的”。基于这个观点,他提出了动物精气理论,认为动物没有心灵,其行为用机器原理就可以解释。动物体内能产生一种叫做“动物精气”(Animal spirits)的物质,能沿着神经管,以类似液压驱动的方式,触发肌肉的活动。
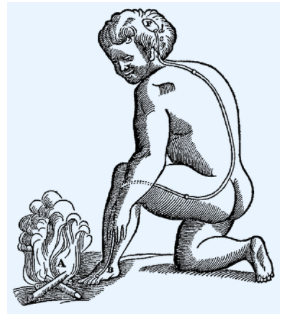
笛卡尔在Treatise of Man中使用的反射路径插图
外界刺激进入到脑,从脑释放出引发肌肉运动的液体,完成反射
这个理论在当时很好地解释了如何产生反射(如上图),但剑桥一位医生弗朗西斯·格里森(Dr. Francis Glisson)却不这么认为,他通过一个简单的实验证实了“动物精气理论”的错误。他让十几个志愿者在水缸中握拳,如果笛卡尔的理论是正确的,肌肉的活动由外部物质(如动物精气)注入而引发,那么在握拳时,外部物质注入使手臂肌肉体积变大,进而水的液面会上升。但在实验结果显示水的液面没什么变化,甚至略有下降,基本宣告了“动物精气理论”的破产,也为后续的研究者留下了谜题——我们究竟如何控制自己的身体。

Dr. Francis Glisson
Jan Swammerdam也做过类似的实验[1]
将青蛙肌肉放在注射器内,刺激裸露的神经触发肌肉活动
同样地,肌肉收缩前后水位没有发生变化
小树分割线
大约百余年后(约1780年),Dr. Luigi Galvani发现用摩擦产生的电荷来刺激青蛙腿的神经末端,会令肌肉收缩变得更强。Galvani称这种控制肌肉的“力量”为动物电,认为动物电是肌肉活动的来源。但当时Galvani并不明白这些“动物电”从哪里来。直到1870年,两位德国医生Dr. Eduard Hitzig 和Dr. Gustave Fritsch才回答了这个问题[2]。

左:Dr. Luigi Galvani;右:其参与的青蛙实验
他们将一只没有麻醉的狗固定,并移除了部分头盖骨,露出大脑,然后用连接在电池上的细铂电极来电击大脑皮质表面。他们发现在电击额叶皮质的某个区域时,会引发肌肉收缩,收缩的程度与电流强度有很强的相关性。随着神经解剖的不断发展,人们认识到该区域为运动皮层。至此,可以回答前面提出的问题,我们躯体的运动受到位于额叶的运动皮层控制。
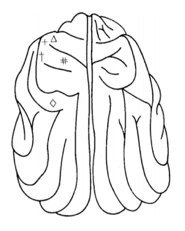
Dr. Eduard Hitzig 和Dr. Gustave Fritsch在狗脑的刺激位置[2]
+ 前腿外展; † 前腿弯曲;
# 前腿运动; ◇ 面部抽搐
那么,运动皮层如何控制我们的身体?
20世纪末,信息论与计算机科学兴起,Dr. Norbert Wiener提出的控制论也深深地影响着心理学与认知科学。在这个技术反哺基础科学的年代,人们把目光投向了由Dr. Bruce Francis及Dr. Walter Wonham所提出的内部模型控制原则(Internal model principle)。
该原则认为可以使用内部模型实现对控制系统的建模,进而能够回答“运动皮层如何控制躯体”的问题。假设躯体是一个控制系统,那大脑对躯体的控制过程可以拆解成若干内部模型的组合。目前,理论神经科学研究者认为,在运动控制中,内模分为前向模型(Forward model)和反向模型(Inverse model)两种。
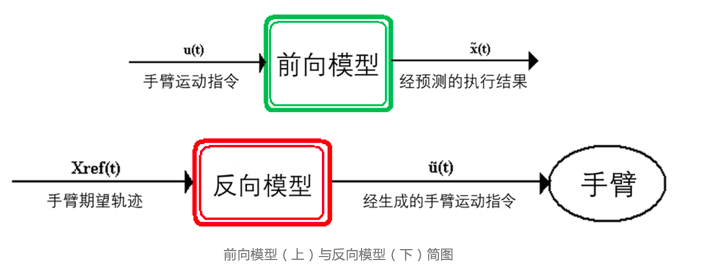
前向模型(上)与反向模型(下)简图
前向模型能够根据运动指令预测运动状态,而反向模型能使用运动状态来推测运动指令。就像一台机械臂,机械臂的伸手方向由运动状态来描述,但伸手行为则与机械臂内部电机对运动指令的执行有关。

“ 革命性假肢”计划:由约翰霍普金斯大学应用物理实验室开发的机械假肢
简单来说,肌肉活动来源于肌单元(Motor unit)的收缩,而肌单元受到来自脊髓的运动神经元支配,这意味着中枢神经系统会使用反向模型将期望的躯体动作转换为给肌单元发送的信号,进而实现对肌肉活动的控制。
但神经脉冲在神经纤维的传递比我们想的慢得多(每秒27米),比如视觉信号到视觉皮层就存在72毫秒的延时[3],如果没有前向模型预测运动状态来进行辅助,我们的运动控制会因感觉信号延时所产生的误差累计而崩溃。
总结来说,我们日常生活中对躯体的灵巧控制是通过这两类内部模型互相配合所实现的。尽管Dr. Daniel Wolpert通过被试者在虚拟视觉反馈系统(Virtual visual feedback system)中的表现巧妙的证明了这一理论[4],但研究者们仍希望在生物体的神经系统中找到前向模型与反向模型的理论基础。
小树分割线
1982年Dr. Apostolos Georgopoulos通过记录猕猴手臂朝向不同方向运动时运动皮层单个神经元的活动发现[5],伸手方向可以通过简单的余弦函数转化为记录到的运动皮层神经元活动,而反过来使用线性模型做加权平均,就能将皮层神经元活动的数据中还原为猕猴手臂活动的轨迹。这一研究很好地为前向模型与反向模型的存在提供了证据支持,并在表达模型中发展出了新的研究方向。
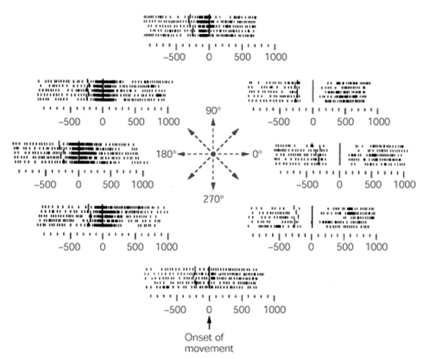
伸手方向不同,神经元的发放频率不同,其发放率存在对伸手方向的偏好[4]
Dr. Georgopoulos认为大部分运动皮层神经元都存在躯体运动状态的偏好方向 (Preferred direction) ,所有神经元的活动偏好就像一个运动指令集储存在运动皮层中,当某一类神经元活动增强时,意味着躯体要执行该类神经元偏好的运动状态。

橘色向量(算法重构的伸手方向)与黄色向量(实际的伸手方向)的方向非常接近[5]
自那以后,对运动皮层偏好方向的研究成为了当时的热点,受偏好方向启发的群体向量算法(Population vector)[6]更是成为日后脑机接口的基石,帮助人类实现了完全的意念控制。可以说,在将运动皮层的运动指令转换为运动状态(伸手方向)也就是前向模型的这个部分,偏好方向的表现非常好。

猕猴通过脑活动控制机械臂完成食物的抓握与进食
小树分割线
然而在2007年斯坦福大学的Dr. Krishna Shenoy指出[7],Dr. Georgopoulos的工作忽略了神经元活动在不同时间尺度的差异,而且测量的伸手行为也较简单,当应用场景比较复杂的时候,Dr. Georgopoulos的偏好方向模型就不再适用。
这戳到了偏好方向模型研究者的痛处:尽管Dr. Apostolos Georgopoulos的偏好方向模型很好的拟合了神经元在整个伸手行为中的平均发放频率,但如果只截取某一小段时间而非整个伸手行为,这个模型的表现就很差,即使给出实际的伸手行为,也没有哪个研究者敢打保票说他能根据已知行为准确推测不同时间的运动皮层活动,实在是当时祖师爷限定的实验条件太单一,苦了后来的继任者。
在2012年Dr. Krishna Shenoy根据群体神经元的记录结果证明了运动皮层的活动具有动态性[8],这意味着引入时间变量的动态模型能比只分析神经元活动与伸手方向之间关系的偏好方向模型更好地解释运动皮层如何编码躯体运动。在后续的工作中,Dr. Shenoy猜想运动皮层是一个由数百万神经元广泛连接组成的递归神经网络(Recurrent neural network, RNN),通过输入和反馈信号与大脑的其他部分连接起来[9]。

动态系统的可视化样例
动力系统在相平面中随时间演化的轨迹
实心圆点代表着该动力系统的吸引子,红色星号为系统的不稳定定点
小树分割线
大脑存在与递归神经网络类似的结构,以一种极端重复的方式连接在一起,此外,RNNs是一种非常通用的计算装置,能用于描述大多数连续时间的动态系统,因此用RNN来对运动皮层建模是一个非常好的选择。但是如何成功地训练RNNs来完成特定的任务是一个需要解决的难题。
不过这个问题在2015年被Dr. Shenoy的博士后Dr. David Sussillo给解决了,Dr. Sussillo是计算神经科学巨擘Dr. Larry Abbott的弟子,在博士期间提出了FORCE learning算法用于RNNs的训练,相较传统的回声状态网络(Echo state network, ESN)和液体状态机(Liquid state machine, LSM)算法,FORCE learning训练的RNNs更加稳定,对多种周期信号的拟合与生成有着不错的表现[10]。

用RNNs生成人的跑步运动[10]
Dr. Sussillo进一步完善了Dr. Shenoy的理论框架,给出了一种重构运动皮层网络的可能(原文用的是“一个解”)[11]——只要对完成训练的RNNs输入指定的运动状态,该网络就会自动输出肌电。更加神奇的是,该网络模拟的神经元活动与实际在猕猴中记录到的神经元活动非常相似,这意味着这个动态模型能够根据运动状态,反向生成运动皮层的神经元活动。虽然该网络能很好地拟合实验数据,但是Dr. Sussillo没有对RNNs网络的权重进行限制,导致网络中有些神经元的发放率无法恢复到0,与实际的电生理数据不符。
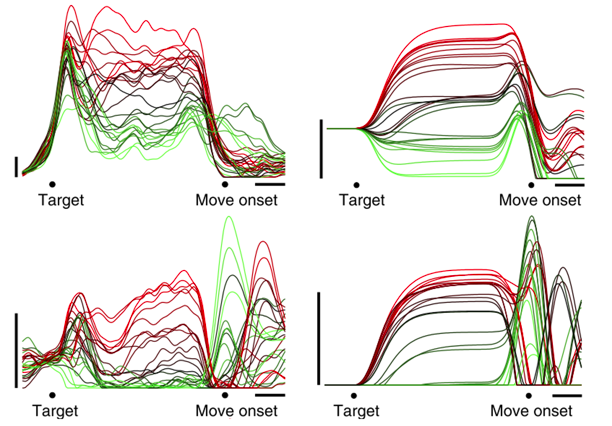
RNNs对运动皮层的重构结果[11]
左侧为实际记录到的神经元活动,右侧为RNNs神经元的活动
顺带一提,Dr. Abbott的徒孙Dr. Guillaume Hennequin在2014年也给出另外的解[12],提出了一种更加仿生的稳态优化环路(Stability-optimized circuits, SOCs),能做到与Dr. Sussillo的工作类似的效果,还能保证网络中的神经元活动在没有外界输入的情况下恢复为0。后续Dr. Jake Stroud又在SOCs上做了些修修补补的工作[13],这里就不细讲了。
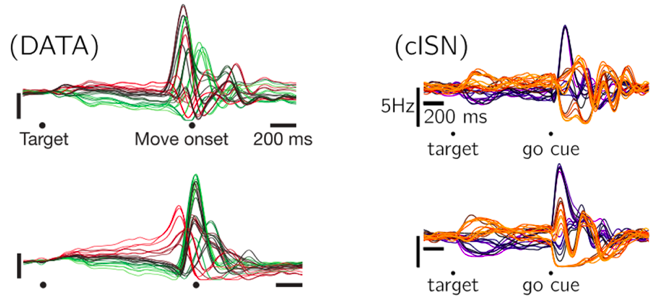
SOCs的重构效果[12]
左侧为记录的运动皮层神经元活动,右侧为SOCs神经元活动
借助数学模型对皮层数据的分析,我们明白运动相关的神经系统存在前向预测运动状态和反向推测运动指令的潜力,在实际的神经生物学研究中也有证明前向模型与反向模型的诸多例子[14],这些结果基本支持了内部模型控制原则,也回应了自笛卡尔提出以来的数百年难题——我们如何控制自己的身体?
郑重声明:本文版权归原作者所有,转载文章仅为传播更多信息之目的,如作者信息标记有误,请第一时间联系我们修改或删除,多谢。



